Communautés fongiques

Contexte
Dans les écosystèmes forestiers Méditerranéens, les communautés fongiques du sol constituent une interface obligatoire entre les autotrophes et le sol. En particulier, les champignons ectomycorhiziens sont associés aux racines des principales essences Méditerranéennes (Pins, Chênes, Châtaignier, etc…), dont ils assurent l’équilibre hydro-minéral, en pourvoyant leur hôte en eau et en formes minéralisées de l’azote et du phosphore d’origine organique.
Sous l’angle de cette diversité, les forêts méditerranéennes constituent un point chaud, particulièrement lorsqu’elles sont dominées par les chênes, un hectare de yeuseraie monospécifique pouvant par exemple receler plusieurs centaines d’espèces (Richard et al., 2004, 2005).
A l’aide d’un suivi effectué sur le dispositif expérimental de Puéchabon, il a été montré que les changements climatiques prévus dans la région méditerranéenne française sont en mesure de modifier la phénologie du développement de l’appareil végétatif fongique (mycélium) dans le sol, ainsi que la composition des communautés ectomycorhiziennes (Richard et al., 2011). Par ailleurs, des travaux s’appuyant sur des données d’herbier ont également montré que la phénologie de la fructification pourrait être sensiblement modifiée par les changements climatiques en cours dans les forêts européennes (Kauserud et al., 2010). Les conséquences sur l’écosystème forestier (dynamique de la végétation) et sur les hôtes (écophysiologie, phénologie de l’allocation minérale et hydrique) de ces changements dans la phénologie du cycle biologique et dans la composition des assemblages fongiques du sol sont encore mal connues et nécessitent la mise en place de suivis sur le long terme.
Au sein de l’extraordinaire diversité fongique Méditerranéenne, l’Amanite des Césars occupe une place toute particulière, en tant qu’espèce à très forte valeur gastronomique (Yun et al. 2004), mais aussi à dimension sociologique, les places à oronges (sites de fructifications) faisant l’objet d’une tradition de transmission intergénérationnelle orale. En outre, cette espèce constitue un excellent candidat pour l’étude de l’impact des changements globaux sur les patrons de diversité, en raison de son autécologie marquée, se caractérisant à la fois par une forte affinité pour le climat méditerranéen, et par une sensibilité à la couverture forestière et au régime de perturbations. De ce fait, cette espèce répond de manière sensible, en termes géographique, au changement climatique en cours dans l’écorégion.
Objectifs
La tâche d’observation communautés fongiques, dirigée au laboratoire CEFE, consiste à étudier la dynamique temporelle à long terme des communautés fongiques du sol dans la forêt méditerranéenne pour interroger deux axes de questionnement :
- A l’échelle des communautés : quelle est la réponse de la biodiversité fongique du sol au changement climatique en cours dans la région méditerranéenne française ? Quelle est la signature à long terme (et son évolution) de l’empreinte anthropique sur les communautés du sol (par ex. places à charbons, sylviculture – question abordée en lien avec les sciences humaines et sociales) ?
- A l’échelle de l’espèce modèle Amanita caesarea : Quel est l’effet du changement climatique en cours sur la phénologie des fructifications et sur le régime de reproduction de cette espèce emblématique, à l’échelle locale ? Quel est l’effet des évolutions climatiques en cours sur la distribution géographique de cette espèce ?
Le dispositif de suivi s’appuie trois sites d’études situés en France Métropolitaine, dont deux sites continentaux et un site insulaire.
Station expérimentale du Fango
Présentation du transect permanent du Pericato (Corse)
En 1994, un transect permanent d’une superficie de 6400 m2 (40 x 160m) a été installé dans la zone centrale de la réserve de biosphère du Fango (classée réserve MAB en 1977), actuellement en cours de classement en réserve biologique intégrale. Ce dispositif de suivi a été établi au sein d’une yeuseraie n’ayant fait l’objet d’aucune intervention sylvicole depuis 1827 au moins (Panaiotis, 1997).
A l’origine, ce dispositif avait vocation d’étudier la dynamique naturelle de la yeuseraie (Mahmedi 1994 ; Panaïotis, 1996), et plus particulièrement le rôle des trouées naturelles dans la régénération naturelle de cette essence (Panaïotis et al., 1997, 1998). Equipé d’un dispositif de quadrats contigus de 100m2 dont les sommets sont matérialisés in situ (Planche 1), le transect permanent du Perticato a été l’objet d’une cartographie exhaustive de sa végétation (arbres, arbustes, semis, recouvrement des différentes strates de végétation (deux étages de chênaie, deux étages de maquis) et d’une analyse de l’empreinte anthropique (places à charbon, anciens bâtis) présente sur le site. En outre, les trouées naturelles présentes sur le site ont été cartographiées et datées.
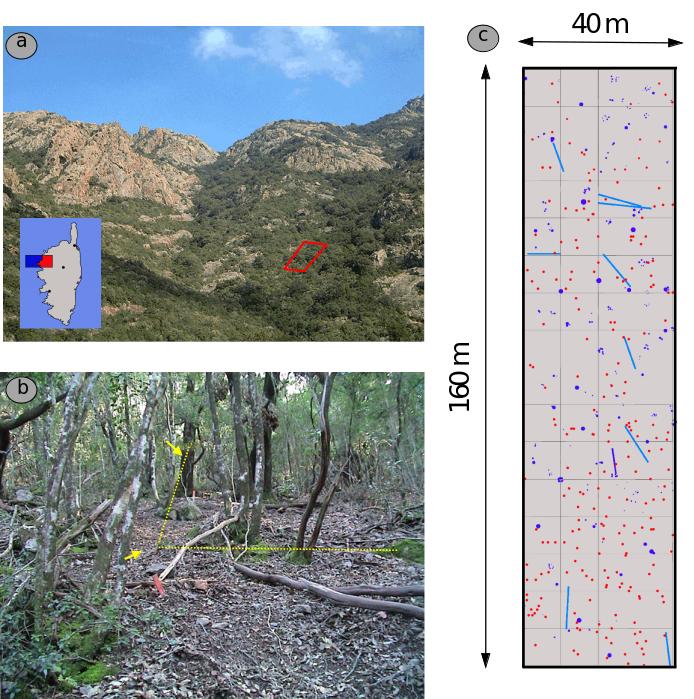
Mesures
Communautés – En 1999, un travail de suivi diachronique des fructifications fongiques a été initié, ainsi qu’une analyse des communautés ectomycorhiziennes du sol. Le suivi diachronique des poussées fongiques porte à la fois sur les communautés (champignons décomposeurs et mycorhiziens) et les populations d’espèces cibles (amanite des Césars). Entre 1999 et 2002, plus de 8000 fructifications de macromycètes ont été cartographiées (Richard et al., 2004).
Population d’amanite des Césars – Les populations d’amanite des césars font l’objet pour leur part d’un suivi continu depuis 1999, avec relevé exhaustif des carpophores (positionnement et prélèvement des carpophores pour génotypage, Planche 2) grâce au soutien apporté par le technicien rivière du Parc Naturel Régional de Corse (signalisation des épisodes de poussées, soutien à la collecte et à la cartographie des carpophores), structure co-gestionnaire de la réserve MAB Fango. Après 16 années de suivi diachronique, 544 carpophores de cette espèce ont été cartographiés, collectés, et analysés à l’aide d’un jeu de 13 marqueurs microsatellites polymorphes développés spécifiquement pour cette espèce au CEFE-CNRS (Salomon, M2 FENEC 2011). L’ensemble des données mycologiques collectées sur le site ont par ailleurs fait l’objet d’une analyse de réseaux d’interactions (Taudière et al., 2015).

Population de Xylobolus subpileatus – Dans le cadre de la thèse d’Adrien Taudière, un relevé exhaustif des fructifications de Xylobolus subpileatus (Planche 3) a été réalisé en 2015. La distribution locale de cette espèce saproxylique, de répartition mondiale associée à une grande rareté, a été étudiée en relation avec l’âge des trouées naturelles qu’il colonise (sur les volis), évaluée avec précision sur tout le transect et dans le massif adjacent (Panaïotis et al., 1998). Ce travail permettra de mieux cerner l’écologie de cette espèce rarissime, spécialisée dans la dégradation des stades les plus avancés de bois mort de chênes. Cette étude a fait l’objet d’une publication en cours d’évaluation (Taudière et al., in review).

Station expérimentale de Puéchabon
Présentation du dispositif permanent de suivi
Le dispositif de suivi des fructifications fongiques s’appuie sur le site de mise en œuvre du programme européen Mediterranean Terrestrial Ecosystems and Increasing installé en 2003 (Planche 4). Ce dispositif consiste en un ensemble de trois blocs expérimentaux séparés de 50 m, comprenant chacun deux quadrats adjacents d’une surface de 140 m2 (14×10 m ; exclusion vs. contrôle).

Mesures
Sur ce site installé en forêt domaniale de Puechabon, une étude des communautés fongiques ectomycorrhiziennes a permis de décrire la diversité des assemblages présents dans la forêt de chêne vert (Richard et al. 2011). Depuis le 12 novembre 2010, un relevé bi-hebdomadaire des fructifications de macromycètes est réalisé, le long d’un transect de 500 mètres linéaires. Les fructifications sont exhaustivement récoltées (J.-M. Ourcival, expérimentateur) et identifiées taxonomiquement au laboratoire CEFE (J.-M. Bellanger, taxonomiste référent). La base de données constituée comprend plus de 600 visites du dispositif, et plus de 100 espèces de macromycètes. Par ailleurs, depuis 2012, le site est équipé de sondes permettant de mesurer en continu la température est l’humidité du sol à 3 et 7 cm de profondeur dans différents points dispositif. Ces mesures permettront de mettre en œuvre prochainement une approche corrélative entre la phénologie des poussées de macromycètes et les conditions environnementales régnant dans le sol.
Réseau de sites de suivi d'Amanite des Césars de Haute-Corrèze
Présentation du dispositif permanent de suivi
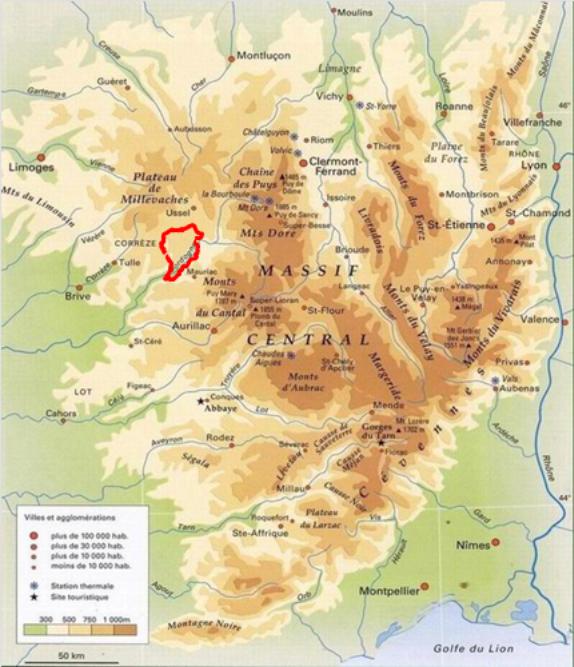
Le dispositif de suivi permanent des Haute-Corrèze a été établi en 2011 afin de compléter le dispositif destiné au suivi de l’amanite des césars dans le cœur de son aire de répartition par un second dispositif d’observation situé en limite continentale de répartition de l’espèce, c’est-à-dire dans son domaine d’expansion sous l’effet des changements climatiques en cours. Cela permettra notamment de comprendre les régimes de reproduction à l’œuvre dans la colonisation de nouveaux sites par cette espèce. Ce dispositif, construit à partir d’une méthode de prospection à l’échelle du territoire d’étude haut-Corrézien (335 km², planche 5), comporte au total 37 stations potentielles, dont 32 se sont révélées productives et sont suivies depuis 2013, qui toutes consistent en un motif répété de pelouse sèche située en bordure de boisements mixtes, au sein de micro-valons de pente moyenne exposés au sud (planche 6).
Mesures
A l’image des relevés effectués sur le site du Fango, les fructifications d’Amanite des Césars sont exhaustivement cartographiées depuis 2012, et les carpophores de cette espèce prélevés pour génotypage par marqueurs microsatellites.
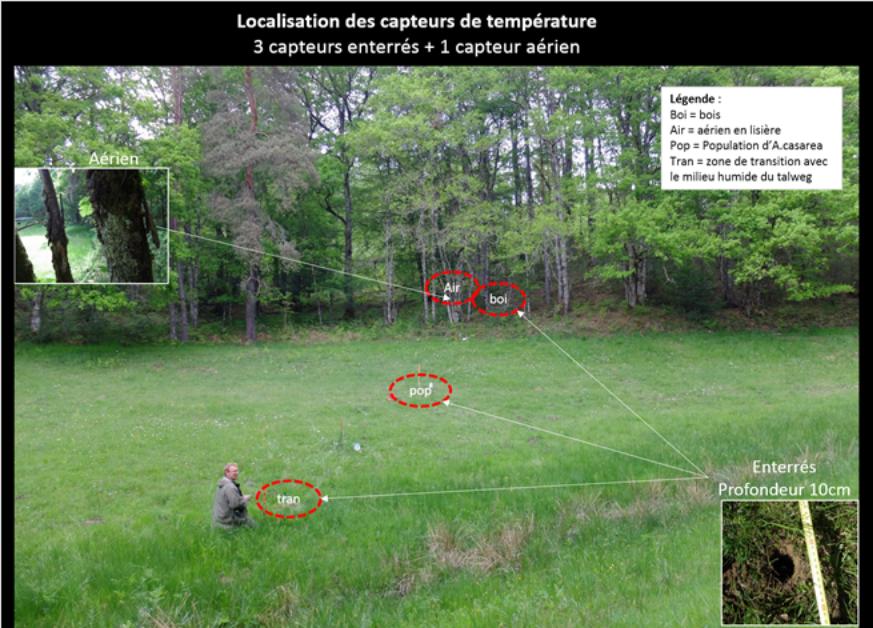
Ces sites ont fait l’objet de descriptions complémentaires de leur biodiversité (communautés fongiques, de végétaux vasculaires, et d’orthoptères), et caractérisés sur le plan microclimatique (stations sont équipées de sondes thermiques de type ibutton, Planche 6), pédologique et sociologiques (historique des pratiques agricoles au cours des dernières décennies). L’ensemble de ces mesures permettra de qualifier l’autécologie de l’amanite des Césars, dans le cadre d’un diplôme de l’Ecole Pratique des Hautes Etudes (Vincent Mennessier).
Données
L’ensemble des données aujourd’hui recueillies se divisent en cinq corpus complémentaires, en lien avec les différentes approches mises en œuvre sur le dispositif de suivi des communautés fongiques.
Herbier de référence d’Amanite des Césars : à l’heure actuelle, cet herbier comporte 967 spécimens, dont 544 spécimens récoltés en Corse, et 423 spécimens échantillonnés en Haute Corrèze.
Base de données génétique de référence d’Amanite des Césars : la totalité des échantillons d’amanite des Césars présents dans l’herbier a fait l’objet d’un génotypage, à l’aide de 13 marqueurs microsatellites polymorphes développés au laboratoire CEFE (échantillons 2016 en cours d’acquisition dans le cadre d’une collaboration Européenne – Short Term Scientific Mission (STSM). COST Action FP1305 – Linking belowground biodiversity and ecosystem function in European forests (BioLink) attribuée à Y. Pinuela Samaniego).
Base de données cartographique des populations de Xylobolus subpileatus : cette base de données contient le positionnement systématique des fructifications de cette espèce dans le site du Fango, soit sur un ensemble de 86 trouées naturelles précisément datées. Cette matrice est associée à une base de données moléculaire (séquence ITS) des carpophores échantillonnés.
Base de données des fructifications de macromycètes du site de Puechabon : cette base de données comporte les relevés de fructifications effectués depuis novembre 2010 sur le transect permanent visité deux fois par semaine. Pour chaque espèce à chaque date de relevé, l’abondance (en nombre de carpophores) est précisée.
Archives de sol : afin de pouvoir réaliser à posteriori des analyses en lien avec les questions posées (dosage d’ADN fongique par qPCR, analyses chimiques, etc…), des archives de sol ont été constituées sur les trois sites de suivi, et stockées au laboratoire CEFE.
Variables Essentielles de Biodiversité (EBV) :
- Species interactions
- Taxonomic diversity
- Allelic diversity
- Population genetic differentiation
- Population abundance
- Species distribution
- Phenology
Portail des données de l'OREME
Page de la TO sur le portail des données de l'OSU OREME
Accéder au portailContact
- Franck Richard (Centre d’Ecologie Fonctionnelle et Evolutive)
- Jean-Michel Bellanger (Centre d’Ecologie Fonctionnelle et Evolutive)
Equipe
Coordinateur : Franck RICHARD (CEFE)
Référents :
- Biologie moléculaire : M.-P. Dubois (CEFE)
- Taxinomie : J.-M. Bellanger (CEFE)
Responsable suivi terrain communautés : J.-M. Ourcival (CEFE)
Etudiants (en cours 2017) : Y. Pinuela Samaniego (COST scholarship), V. Mennessier (EPHE, co-encadrement C. Carcaillet).
Collaborations
Concernant le volet communautés, la tâche d’observation génère depuis sa création (2012) une collaboration transversale au CEFE entre deux départements de recherche (Ecologie Fonctionnelle et Interactions, Ecologie et Sociétés) impliquant à la fois expertise taxonomique et écologique et ingénierie. Ce volet fera prochainement l’objet d’une valorisation sous forme de publication.
Ce volet a par ailleurs permis l’organisation de l’Annual Overseas Foray de la British Mycological Society en Corse en 2014, et faire la première découverte sur le site de suivi du Fango de Xylobolus subpileatus, point de départ d’une collaboration avec le Natural History Museum of Denmark (T. Læssøe).
Concernant le volet populations, la tâche d’observation génère depuis sa création une collaboration étroite avec l’Ecole Pratique des Hautes Etudes, finalisée par le Diplôme EPHE en cours de Vincent Mennesssier. Par ailleurs, ce volet a permis de mettre en place une collaboration transversale au CEFE (Service des Marqueurs Génétiques – M.-P. Dubois) et conduit au co-encadrement de plusieurs étudiants de Licence (A. Christophe, 2015, C. Pillard, 2016) et de Master (Y. Pinuela Samaniego, 2016 et 2017) en agrégeant des financements externes (STSM).
Valorisation scientifique
Volet communautés
Taudiere, A., Munoz, F., Lesne, A., Monnet, A. C., Bellanger, J. M., Selosse, M. A., Moreau, P.-A. & Richard, F. (2015). Beyond ectomycorrhizal bipartite networks: projected networks demonstrate contrasted patterns between early-and late-successional plants in Corsica. Frontiers in plant science, 6.
Volet populations
M.-P. Dubois, V. Salomon, C. Damon, M. Gardes, M.-A. Selosse, H. Gryta & F. Richard 2012. Dynamique des populations d’amanite des Césars (Amanita caesarea) en Corse. 9èmes Rencontres de Phytopathologie – Mycologie de la Société Française de Phytopathologie, Aussois, 16-20 janvier (Poster, International).
F. Richard 2014. Réponse de la diversité fongique au changement climatique : quelles leçons en région méditerranéenne ? Micosylva International Workshop, Poblet (Spain), 26-30 janvier (Oral Session, International).
F. Richard 2015. Linking fungal diversity to the dynamics of Mediterranean ecosystems: lessons from two contrasted case studies. 1st Mediterranean Mycological Congress, Platres (Cyprus), 27-29 novembre (Oral session, International, invited).
Taudiere, A., Moreau, P.-A., Carcaillet, C., Christophe, A., Læssøe, T., Panaïotis, C., Bellanger, J.-M. & Richard, F. (in review). Xylobolus subpileatus, a hyper-specialized basidiomycete functionally linked to old canopy gaps.
Références
Kauserud, H., Heegaard, E., Semenov, M. A., Boddy, L., Halvorsen, R., Stige, L. C & Stenseth, N. C. (2010). Climate change and spring-fruiting fungi. Proceedings of the Royal Society of London B: Biological Sciences, 277(1685), 1169-1177.
Panaïotis, C., Carcaillet, C., & M’Hamedi, M. (1997). Determination of the natural mortality age of an holm oak (Quercus ilex L.) stand in Corsica (Mediterranean Island). Acta Oecologica, 18(5), 519-530.
Panaïotis, C., Loisel, R., & Roux, M. (1998). Analyse de la réponse de la végétation aux trouées naturelles dans une futaie âgée de Quercus ilex L. en Corse (île Méditerranéenne). Canadian journal of forest research, 28(8), 1125-1134.
Richard, F., Moreau, P. A., Selosse, M. A., & Gardes, M. (2004). Diversity and fruiting patterns of ectomycorrhizal and saprobic fungi in an old-growth Mediterranean forest dominated by Quercus ilex L. Canadian Journal of Botany, 82(12), 1711-1729.
Richard, F., Millot, S., Gardes, M., & Selosse, M. A. (2005). Diversity and specificity of ectomycorrhizal fungi retrieved from an old‐growth Mediterranean forest dominated by Quercus ilex. New Phytologist, 166(3), 1011-1023.
Richard, F., Roy, M., Shahin, O., Sthultz, C., Duchemin, M., Joffre, R., & Selosse, M. A. (2011). Ectomycorrhizal communities in a Mediterranean forest ecosystem dominated by Quercus ilex: seasonal dynamics and response to drought in the surface organic horizon. Annals of Forest Science, 68(1), 57-68.
Taudiere, A., Munoz, F., Lesne, A., Monnet, A. C., Bellanger, J. M., Selosse, M. A., Moreau, P.-A. & Richard, F. (2015). Beyond ectomycorrhizal bipartite networks: projected networks demonstrate contrasted patterns between early-and late-successional plants in Corsica. Frontiers in plant science, 6.
Taudiere, A., Moreau, P.-A., Carcaillet, C., Christophe, A., Læssøe, T., Panaïotis, C., Bellanger, J.-M. & Richard, F. (in review). Xylobolus subpileatus, a hyper-specialized basidiomycete functionally linked to old canopy gaps.
Yun, W., & Hall, I. R. (2004). Edible ectomycorrhizal mushrooms: challenges and achievements. Canadian Journal of Botany, 82(8), 1063-1073.